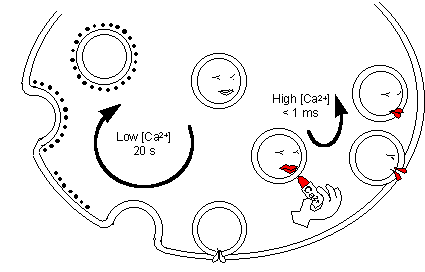
The kiss-and-run model
of neurotransmitter secretion
The clear synaptic vesicles of neurons release their content at the presynaptic
membrane and are then quickly retrieved.
It is unclear whether a complete cycle of exocytosis and endocytosis
is always involved [Heuser and Reese (1973) J. Cell Biol. 57, 315]
or whether neurotransmitter can be released by a transient interaction
[Ceccarelli, Hurlbut and Mauro (1973) J. Cell
Biol. 57, 499  ].
].
Findings in chromaffin and mast cells suggest that exocytosis is preceded
by the formation of a pore that has similar conductance properties to ion
channels. The content of the secretory organelle partially escapes at this
early step, but the pore can close before the vesicle fuses fully [Chow,
Von Ruden and Neher (1992) Nature 356,60; Alvarez de Toledo, Fernandez-Chacon
and Fernandez (1993) Nature 363, 554]
May quantal release of neurotransmitter from clear synaptic vesicles
occur by a similar 'kiss and-run' mechanism? [Neher (1993) Nature
363,497; Fesce, Grohovaz, Valtorta and Meldolesi
(1994) Trends in Cell Biol. 4, 1].
Klingauf et al. [Nature 394, 381, 1998] detected at least two
components of recycling in hippocampal neurons: a slow process (20-30 s)
and a much faster process, which was interpreted as kiss-and-run recycling,
the role of the latter process greatly increasing with stimulus strength
and elevated [Ca2+]. And recently, Alés and colleagues
[Nature Cell Biology 1, 40,1999], by means of cell-attached capacitance
measurements coupled to catecholamine assay by a carbon fibre positioned
within the pipette, showed that in chromaffin cells the full vesicle content
is discharged through reversible fusion. Furthermore, the frequency of
kiss-and-run fusions rises from 5% to almost 80% as calcium concentration
increases, and the duration of the transient opening falls to a few tens
of ms.
Thus, the cells appear to have the choice among
at least two processes for membrane recycling: slow conventional endocytosis
predominates when [Ca2+] is low, fast kiss-and-run when it is
high. If a chromaffin granule, which has a volume about 10 times as large
and is filled with an organised macro-molecular matrix trapping the biogenic
amines, is fully discharged in few ms, it is well possible that a synaptic
vesicle releases its soluble neurotransmitter in a fraction of a millisecond
and rapidly pinches off from the presynaptic membrane [Fesce and Meldolesi
(1999) Nature Cell Biology 1, E3].

Local recycling of synaptic vesicles
first proposed by B. Ceccarelli et al.
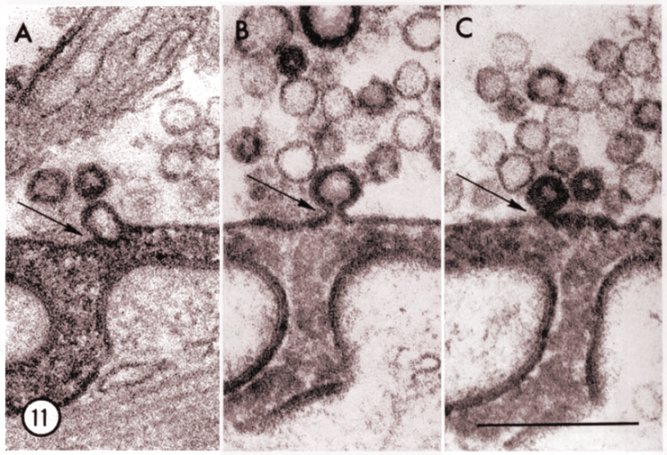
High power electron micrograph of portions of three different neuromuscular
junctions at the level of the "active zone". The preparations were stimulated
for 2 hr at 2/sec in curare plus horseradish peroxidase. Junctional clefts
contain rich deposits of peroxidase reaction products. The figures show
three degrees of association between peroxidase-labeled vesicles and the
prejunctional membrane (arrows). In A the membrane of the vesicle is completely
fused with the prejunctional membrane; in B the continuity of the two membranes
is mantained through a short stalk; and in C the vesicle appears to be
in the process of losing contact with the axolemma. [calibration
bar: 0.25 mm]
-
BRUNO CECCARELLI, W.P.
Hurlbut and A. Mauro.
Depletion of vesicles from frog neuromuscular junctions by prolonged
tetanic stimulation.
Journal of. Cell Biology 54, 30-38, 1972.
Vesicle fusions frozen during transmitter
release:
fusion pores can be seen, wide-open vesicles are rare,
late events
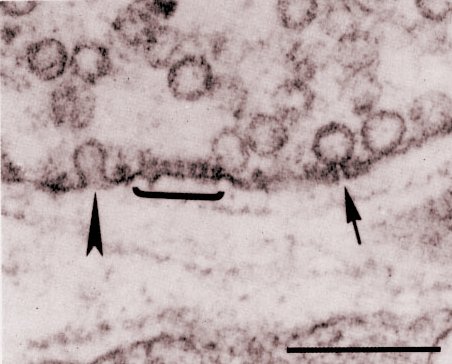
Electron micrograph of a cross section from a neuromuscular preparation
quick-frozen 10 ms after a single stimulus in the presence of 4-aminopyridine
(which prolongs the time course of the evoked release) and processed as
described in [Torri Tarelli, Grohovaz, Fesce and Ceccarelli (1985) J.
Cell Biol. 101,1386].
The bracket highlights the structure of an active zone, facing the post-junctional
infolding.
The arrow indicates a vesicle open to the synaptic cleft through a narrow
pore.
The arrow head indicates a wide-open vesicle fusion (omega shape)
as could occasionally be seen only at late times (>5 ms)
The kiss-and-run model
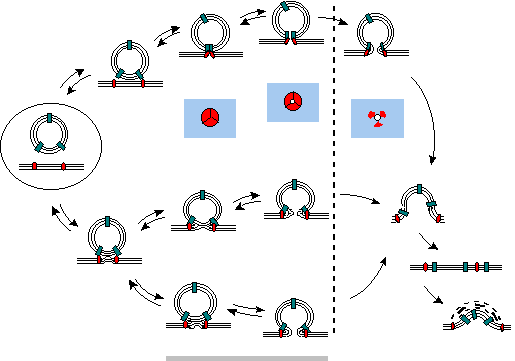 Diagrammatic representation of three possible pathways for vesicle fusion
after the vesicle has docked with to the presynaptic membrane
top: Protein-protein interactions mediate the formation of a supramolecular
complex that operates as the reversible fusion pore. upon widening
of the pore the complex disassembles by the inclusion of mobile lipids,
and full fusion occurs
middle: A scaffold of proteins produces a local perturbation of
the bilayers that leads to hemifusion; a reversible pore opens across the
the resulting single bilayer and eventually leads to complete fusion
bottom: The classical apposition-fusion-fission sequence through
the formation of a pentalaminar structure.
Diagrammatic representation of three possible pathways for vesicle fusion
after the vesicle has docked with to the presynaptic membrane
top: Protein-protein interactions mediate the formation of a supramolecular
complex that operates as the reversible fusion pore. upon widening
of the pore the complex disassembles by the inclusion of mobile lipids,
and full fusion occurs
middle: A scaffold of proteins produces a local perturbation of
the bilayers that leads to hemifusion; a reversible pore opens across the
the resulting single bilayer and eventually leads to complete fusion
bottom: The classical apposition-fusion-fission sequence through
the formation of a pentalaminar structure.
In all cases the fusion is followed by collapse of the vesicle membrane
into the axolemma, intermixing of the component, and subsequent sorting
and retrieval by coated pits and vesicles.
The gray bar highlights the reversible opening that may account for kiss-and-run
release.
-
Riccardo Fesce, Fabio Grohovaz, Flavia Valtorta and Jacopo Meldolesi.
Neurotransmitter release: fusion or 'kiss-and-run' ?
Trends in Cell Biology 4, 1-4, 1994.
-
Riccardo Fesce and Jacopo Meldolesi.
Peeping at the vesicle kiss
Nature Cell Biology 1, E3-E4, 1999.
Bruno Ceccarelli Association
R. Fesce homepage